






Lo más simple es explicar este fenómeno por una única fusión de dos cromosomas en el linaje humano. ¿Pero qué evidencias hay de ello?

En los últimos meses han aparecido en las páginas de Protestante Digital varios artículos cuestionando resultados ya antiguos que relacionan genéticamente a los humanos y los grandes simios. No solamente se ponen en duda estos resultados (muy numerosos y publicados en diferentes revistas especializadas de diferentes países desde hace casi medio siglo) sino que se aprovecha la ocasión para cuestionar la honestidad de los científicos, algo a lo que los autores creacionistas de la tierra joven (CTJ) y vieja (CTV) nos tienen ya acostumbrados cada vez que se publica algo que no encaja con sus ideas1.
Tal vez las críticas más llamativas tienen que ver con dos temas muy conocidos que se usan habitualmente para mostrar la existencia de un antepasado común entre humanos, chimpancés y otros grandes simios: (1) la fusión de dos cromosomas ancestrales para formar el actual cromosoma 2 humano y (2) la existencia de cientos de miles de elementos móviles compartidos entre los genomas de humanos, chimpancés y otros grandes simios. Dado que a esto último dediqué varios artículos en un debate en 2011 en estas páginas2, vamos a centrarnos aquí en el primer punto.
CROMOSOMAS: HUMANOS 23 - CHIMPANCÉS 24
Las publicaciones iniciales del CTJ/CTV, bastante toscas, han dado paso en los últimos años a textos más sofisticados que, usando una jerga científica a veces deliberadamente oscura, pretenden elevar el nivel de la discusión a unos tecnicismos que solo alguien con ciertos conocimientos científicos puede seguir. Han creado incluso sus propias revistas de apariencia científica y con todo ello pretenden, y con cierta frecuencia consiguen, impresionar, especialmente al público evangélico. Algunas de sus publicaciones (que han acabado por tener eco en Protestante Digital) niegan que el cromosoma 2 sea el resultado de la fusión de dos cromosomas ancestrales3.
Un cromosoma es uno de los segmentos en los que se divide toda la información genética de una célula (el genoma). En el caso humano se trata de 23 cromosomas, mientras que chimpancés, gorilas y orangutanes tienen sus genomas, que son de similar tamaño y composición, divididos en 24 cromosomas. ¿Por qué poseemos uno menos si según plantea la evolución tenemos antepasados comunes? Claramente lo más simple es explicar este fenómeno por una única fusión de dos cromosomas en el linaje humano (frente a divisiones en todos los demás grandes simios). ¿Pero qué evidencias hay de ello?
A principios de los años 70 del siglo XX aparecieron diversas publicaciones de autores franceses (como C. Turleau y J. de Grouchy) o españoles (J. Egozcue) que utilizaron sustancias químicas para teñir cromosomas de humanos y chimpancés, y luego gorila y orangután. Estas tinciones son más o menos intensas dependiendo de la composición química del ADN que forma los cromosomas, generando un patrón de bandas a lo largo del cromosoma que recuerda un código de barras y que permite comparar los cromosomas de una especie con otra. Por si alguien está preocupado, no se trata de oscuras técnicas ‘evolucionistas’ o ‘darwinistas’, sino técnicas usadas rutinariamente en el estudio de los cromosomas de diferentes especies animales y vegetales. Técnicas que también permiten detectar alteraciones cromosómicos, comparando individuos dentro de una misma especie, como un Síndrome de Down debido a la aparición un cromosoma adicional, que por estos métodos se comprueba que corresponde al cromosoma 21.
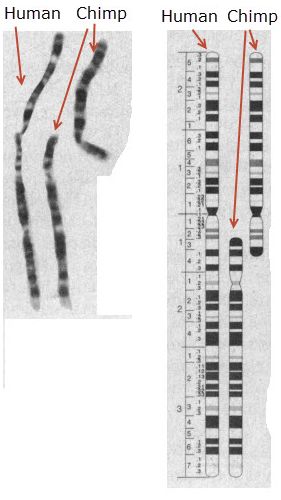
Estos estudios fueron seguidos por los de J.J. Yunis en E.E.U.U., a principios de los años 80, que consiguieron comprobarlos con mucho más detalle, obteniendo patrones muy finos de bandas en los cromosomas de las cuatro especies que permitían comprobar claramente que la composición de todos sus cromosomas era muy similar (véase la imagen comparativa)4. Otras técnicas confirmaron posteriormente estos resultados, como la “Fluorescence in situ hybridization” (FISH)5. Por la imagen anterior quedaba claro que el cromosoma 2 humano correspondía al alineamiento de dos cromosomas de grandes simios. Toda su arquitectura tenía un equivalente en la suma de otros dos cromosomas, apuntando a que el cromosoma 2 humano era el resultado de la fusión de dos cromosomas ancestrales que, sin embargo, permanecieron separados en los linajes de chimpancés y otros grandes simios.
La disponibilidad de técnicas de secuenciación desde finales de los años 70 del siglo XX permitió leer directamente la información del ADN, examinando la sucesión exacta de las cuatro moléculas que componen a manera de ladrillos esta cadena y que se denominan por las iniciales de sus nombres (A,T,C y G). Finalmente, a principios de este siglo, fue posible obtener las secuencias completas de los genomas de humanos y grandes simios. Esto permitió comparar toda la información contenida en el ADN al mayor nivel posible de detalle (sus moléculas químicas) y ver que, como se esperaba desde principios de los años 70 en base a las técnicas de tinción, la similitud era enorme entre aquellas cuatro especies de primates: cerca del 98% (los valores exactos pueden variar dependiendo de las diferentes formas de calcular la similitud, pero muy poco, a pesar de los desesperados intentos de los CTJ/CTV por buscar ahí un motivo de crítica a la evolución)6.
¿UNA FUSIÓN DE CROMOSOMAS EN EL CROMOSOMA 2 HUMANO?
La posibilidad de secuenciar el ADN del cromosoma 2 humano ha permitido localizar y estudiar en detalle el punto exacto de lo que se había propuesto ya en los años 70 como una fusión de dos cromosomas ancestrales. Los cromosomas son segmentos lineales de la doble cadena de ADN, por lo que la secuencia de información en sus extremos (telómeros) es especial para evitar que se deteriore (evitar una especie de ‘deshilache’). Se trata de repeticiones de la secuencia TTAGGG. Por otro lado, los cromosomas tienen una secuencia especial en alguna zona de su interior que dirige la separación de las dos copias cromosómicas durante la división celular. Son regiones conocidas como centrómeros. Pues bien, la fusión de cromosomas permite predecir que se encontrarán dos telómeros enfrentados a ambos lados del punto de fusión, y que el cromosoma fusionado resultante tendrá dos centrómeros (uno seguirá activo y el otro se predice que esté inactivo y no sea funcional, véase figura adjunta).

Todo esto se comprobó a partir de la década de los 90, confirmando lo que ya se había propuesto veinte años antes en base a las tinciones cromosómicas mediante el análisis de la secuencia de la región del cromosoma 2 correspondiente al punto exacto de la fusión7, y volvió a confirmarse con la secuenciación completa del genoma humano a principios de este siglo8, así como el de neandertales y denisovanos que también reflejan una fusión en su cromosoma 29. De hecho, es posible vivir con importantes reorganizaciones en los cromosomas10, e incluso se han descrito personas perfectamente sanas con cromosomas fusionados, como un caso reciente con 22 cromosomas debido a la fusión de los cromosomas 14 y 1511.
¿Y entonces a qué vienen las críticas recientes de los CTJ/CTV12? Básicamente se centran en la identificación del punto de fusión y el centrómero inactivado13.
¿PROBLEMAS EN EL PUNTO DE FUSIÓN?
Se argumenta que el número de repeticiones de la secuencia telomérica TTAGGG es muy inferior al que se supone teniendo en cuenta los números de repeticiones en los telómeros intactos de los cromosomas (de varios cientos o unos pocos miles frente a unas decenas). Por otro lado se argumenta la existencia de secuencias teloméricas internas repartidas por todo el genoma y que no se asocian a un evento de fusión. Ambas cosas han sido explicadas en la bibliografía científica relevante.
Brevemente, aunque un evento de fusión cromosómica es algo excepcional, puede recrearse en condiciones de laboratorio donde se asocia a una gran inestabilidad de los telómeros, que pierden gran número de sus repeticiones antes de poder llegar a fusionarse14. Esta situación de degradación de las repeticiones teloméricas en el punto de fusión no es, por tanto, una evidencia contraria el hecho de haber habido una fusión. Si a esto añadimos la gran antigüedad de este evento de fusión, no deberíamos sorprendernos que la ‘cicatriz’ resultante de la fusión haya borrado en gran medida la secuencia de los extremos originales de los cromosomas fusionados. Esto es más que una especulación, dado que las repeticiones invertidas constituyen zonas muy inestables: véase Ijdo y col. (1991) para los detalles y referencias relevantes. De hecho, una parte de las repeticiones teloméricas TTAGGG son imperfectas debido a pequeñas inserciones, deleciones y mutaciones.
Aunque es un tema de estudio desde los años 90, algunos CTJ/CTV mencionan, como si fuera un secreto oculto, que las secuencias teloméricas no son exclusivas de los telómeros, y que hay repeticiones teloméricas en varias regiones internas de los cromosomas (secuencias intersticiales teloméricas, interstitial telomeric sequences, ITSs). Se supone que así se invalida que el hallazgo de los dos telómeros enfrentados sea una evidencia en favor de una fusión de cromosomas ancestrales en el cromosoma 2. En ese contexto, la referencia frecuente a un trabajo de investigadores liderados por la bióloga Montserrat Ponsà de Barcelona carece de sentido15, dado que ni su artículo ni ella misma apoyan las interpretaciones de los CTJ/CTV. En ese artículo se indica que hay al menos tres tipos de ITSs, las derivadas de una fusión de cromosomas (“fusion ITSs”), las derivadas de una recombinación en los telómeros (“subtelomeric ITSs”) y las derivadas de una reparación de rupturas de doble banda de ADN (“short ITSs”). Precisamente el caso del cromosoma 2 se señala en ese artículo como un caso bien estudiado de fusión de cromosomas, en el que se encuentran dos regiones teloméricas enfrentadas en sentidos opuestos, y se da como referencia el artículo de Ijdo y col. (1991). Pero no todas las ITSs tienen que ver con eventos de fusión de cromosomas, y el artículo del grupo de Ponsà se centra en estudiar inversiones de fragmentos de cromosomas (en los que se produce un corte y también una fusión de fragmentos de ADN).
MÁS ALLÁ DEL PUNTO DE FUSIÓN...
Es más, si nos tomamos la molestia de leer el artículo de Ijdo y col. (1991) podemos encontrar que no solamente se describe la secuencia exacta del punto de fusión con un buen número de repeticiones enfrentadas frente a frente a ambos lados (varias decenas). Los telómeros son algo más que unas repeticiones monótonas de la secuencia TTAGGG. Un telómero tiene también una secuencia pre-telomérica, antes de llegar al extremo del cromosoma. Y, como era de esperar, en el punto de fusión del cromosoma 2 no solamente hay dos telómeros fusionados frente a frente, sino que por fuera de ellos hay dos secuencias pre-teloméricas, una a cada lado: véase la imagen original en la Fig. 1B de Ijdo y col. (1991) y aquí una versión más clara en colores16. Así que no se puede decir que el punto de fusión no esté claramente identificado. Mirar tanto el punto exacto de fusión hace que algunos no vean ciertos detalles relevantes en los alrededores…

Y otro de esos detalles es que tanto humanos como chimpancés comparten una inversión de una pequeña región que rodea al centrómero activo del cromosoma 2 humano (y su equivalente en chimpancés). Esa inversión no se encuentra en gorilas y orangutanes, por lo que podemos concluir que, tras la separación de gorilas y orangutanes, la inversión tuvo lugar en el antepasado común de humanos y chimpancés. Tras su posterior bifurcación, y tan solo en el linaje humano, se produjo la fusión de cromosomas (véase figura adjunta)17.
Pero lo que más llama la atención es que a algunos los ‘árboles’ teloméricos no les permitan ver el ‘bosque’ cromosómico. Mientras discuten si el punto de fusión está más o menos claramente preservado, suponen que una falta de claridad ahí invalida la fusión cromosómica, diciendo que toda la evidencia para la fusión de cromosomas está en la existencia de esa secuencia puntual de fusión. Resulta fascinante que no se den cuenta que es todo el cromosoma 2 entero, y su paralelo aplastante con otros dos cromosomas de chimpancé (2A y 2B), lo que muestra a las claras que ahí ha habido una fusión, como ya se podía ver en las imágenes de Yunis y Prakash en 1982. Es como juntar dos libros de millones de páginas y que luego alguien ponga en duda la fusión porque faltan algunas páginas en la juntura. Por si fuera poco esto, la existencia de un segundo centrómero inactivo se encontró en el lado esperado de la fusión (el 2q)18. Incluso aunque en el punto de fusión se hubieran borrado por completo las regiones teloméricas terminales de los cromosomas originales y el segundo centrómero, la fusión seguiría siendo evidente a lo largo de todo el cromosoma 2 humano y los cromosomas 2A y 2B de chimpancé (y sus equivalentes en otros grandes simios). ¡Solamente hay que mirar la figura 2 de Yunis y Prakash de 1982!19
¿Y QUÉ HACEMOS DISCUTIENDO ESTO AQUÍ?
De un tiempo a esta parte me pregunto esto cada vez que veo artículos negando diferentes porciones del conocimiento científico en medios evangélicos de difusión general: ya sea negando la evolución, el Big Bang, el movimiento de la tierra, o ciertos aspectos del estudio literario o lingüístico de los textos bíblicos.
Me pregunto qué ganan algunos enturbiando ciertos campos de estudio con la pretensión de ‘defender’ a Dios. Además de desorientar a los lectores sin un trasfondo científico y hacer ‘perder’ mucho tiempo a los científicos que tenemos que refutar semejantes argumentaciones, poco más se consigue al final. Lo único que se está haciendo es defender una determinada forma muy particular de entender la fe y las relaciones ciencia y fe que parece necesitar la confrontación con ciertas ideas científicas establecidas. Aunque a veces se presente esto como concebido con la buena intención de defender la ‘verdadera ciencia’, me pregunto por qué los CTJ/CTV no nos obsequian con su actividad en pro de la ciencia en otros campos científicos novedosos que no tengan implicaciones doctrinales de su interés.
Me temo que se trata de una estrategia para hacer una ‘apologética’ muy dudosa y de alto riesgo porque promueve la idea de un conflicto entre el cristianismo y la ciencia contemporánea. Pero esa forma de entender el cristianismo no es la única que existe y los cristianos no necesitamos enfrascarnos en estas polémicas con los científicos. A Dios no hay que defenderlo así porque como decía C.S. Lewis, “una ciencia retorcida en interés de la apologética sería un pecado y una locura.”20
Un cristiano puede admirar la creación y adorar a su Creador independientemente de lo que pase en el cromosoma 2.
Agradecimientos: esta publicación ha sido posible gracias al apoyo de una donación de la Templeton World Charity Foundation, Inc. Las opiniones expresadas en esta publicación pertenecen al autor y no reflejan necesariamente los puntos de vista de la fundación.
Notas
1# Cruz, A. ¿Es el cromosoma 2 humano la unión de dos cromosomas del chimpancé? Protestante Digital, 14 de enero de 2017. Hombres, simios y retrovirus. Protestante Digital, 5 de febrero de 2017. Las ardillas voladoras y la teoría del ancestro común. Protestante Digital, 11 de febrero de 2017.
2# Por orden cronológico, el debate se desarrolló así:
Antonio Cruz. La inquisición darwinista, 16 enero 2011: http://protestantedigital.com/magacin/10179/La_inquisicion_darwinista
Pablo de Felipe. ¿La inquisición darwinista?, 20 marzo 2011: http://protestantedigital.com/magacin/11741/iquestLa_inquisicion_darwinista
Antonio Cruz. El truco darwinista de los genes saltarines, 10 de abril 2011: http://protestantedigital.com/magacin/11794/El_truco_darwinista_de_los_genes_saltarines
Pablo de Felipe. No hay truco con los genes saltarines, 17 de abril 2011:
http://protestantedigital.com/magacin/11804/No_hay_truco_con_los_genes_saltarines
Antonio Cruz. La nueva genética contradice a Darwin, 15 de mayo 2011: http://protestantedigital.com/magacin/11863/La_nueva_Genetica_contradice_a_Darwin
Pablo de Felipe. La nueva genética confirma la evolución, 22 mayo 2011:
http://protestantedigital.com/magacin/11878/La_nueva_genetica_confirma_la_evolucion
3# Bergman, J. y Tomkins, J. The chromosome 2 fusion model of human evolution-part 1: re-evaluating the evidence. Journal of Creation 25 (2011):106-110. Tomkins, J. y Bergman, J. The chromosome 2 fusion model of human evolution-part 2: re-analysis of the genomic data. Journal of Creation 25 (2011):111-117.
4# Yunis, J.J., Sawyer, J.R. y Dunham, K. The Striking Resemblance of Hihg-Resolution G-Banded Chromosomes of Man and chimpanzee. Science 208 (1980):1145-1148. Yunis, J.J. y Prakash, O. The Origin of Man: A Chromosomal Pictorial Legacy. Science 215 (1982):1525-1530.
5# Wienberg, J. y col. The origin of human chromosome 2 analyzed by comparative chromosome mapping with a DNA microlibrary. Chromosome Research 2 (1994):405-410. Kasai, F. y col. Comparative FISH mapping of the ancestral fusion point of human chromosome 2. Chromosome Research 8 (2000):727–735.
6# Se considera que la similitud está entre un 96% y 99%. Varki, A. y Altheide, T.K. Comparing the human and chimpanzee genomes: Searching for needles in a haystack. Genome Res. 15 (2005):1746-1758.
7#Ijdo, J.W. y col. Origin of human chromosome 2: An ancestral telomere-telomere fusion. Proc. Natl. Acad. Sci. USA 88 (1991):9051-9055.
8# Hillier, L.W. y col. Generation and annotation of the DNA sequences of human chromosomes 2 and 4. Nature 434 (2005):724-731.
9# Meyer, M. A high-coverage genome sequence from an archaic Denisovan individual. Science 338 (2012):222–226.
10# O’Neill, I. D. Homozygosity for constitutional chromosomal rearrangements: a systematic review with reference to origin, ascertainment and phenotype. Journal of Human Genetics 55 (2010):559–564.
11# Wang, B. y col. Case Report: Potential Speciation in Humans Involving Robertsonian Translocations. Biomedical Research 24 (2013):171-174. Song, J. y col. A family with Robertsonian translocation: a potential mechanism of speciation in humans. Molecular Cytogenetics 9 (2016):48. Another case is described by Miryounesi, M. y col. Homozygosity for a Robertsonian Translocation (13q;14q) in a Phenotypically Normal 44, XX Female with a History of Recurrent Abortion and a Normal Pregnancy Outcome. J. Reprod. Infertil. 17 (2016): 184-187.
12# Curiosamente, Fazale Rana, uno de los líderes de una de las organizaciones principales del CTV, Reasons to Believe, parece aceptar un evento de fusión en la formación del cromosoma 2 humano, aunque lo considera algo imposible explicable solamente por la acción de Dios. Rana, F. Chromosome 2: The Best Evidence for Evolution? Reasons to Believe, 1 de junio de 2010.
13# Otras críticas, como la presunta existencia de un gen con parte a un lado y parte a otro del punto de fusión, requieren de un espacio más amplio para su análisis del que corresponde a estos artículos y de una profundización mayor en Biología Molecular. Esta idea fue lanzada en otra revista CTJ por Tomkins, J.P. Alleged Human Chromosome 2 “Fusion Site” Encodes an Active DNA Binding Domain Inside a Complex and Highly Expressed Gene—Negating Fusion. Answers Research Journal 6 (2013):367–375. Ha sido criticada con mucho detalle en http://theleagueofreason.co.uk/viewtopic.php?f=8&t=12424 y en otros de los blogs y foros mencionados en la nota al pie número 19.
14# Blasco, M.A. y col. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell 91 (1997):25-34.
15# Farré, M. Interstitial telomeric sequences (ITSs) are not located at the exact evolutionary breakpoins in primates. Cytogenet. Genome Res. 124 (2009):128-131.
16# Los estudios de Ijdo y col. (1991) fueron confirmados en gran detalle una década más tarde en la estela de la secuenciación del genoma humano: Fan, Y. y col. Genomic Structure and Evolution of the Ancestral Chromosome Fusion Site in 2q13-2q14.1 and Paralogous Regions on Other Human Chromosomes. Genome Research 12 (2002):1651-1662.
17# Tomado de Venema, D. Los genomas como textos antiguos, parte 5. Curso Fundamentos de la evolución, lección 19 (publicado originalmente en la página web de BioLogos Foundation, 18 de julio de 2013, y traducido por el Centro de Ciencia y Fe).
18# Avarello, Ri y col. Evidence for an ancestral alphoid domain on the long arm of human chromosome 2. Hum Genet 89 (1992):247-249. Véase también la referencia de Fan y col. 2002 en la anterior nota número 16.
19# Para una discusión más detallada y técnica, pueden verse diversos blogs y foros que han analizado los artículos citados de Tomkins y Bergman:
http://bioinformaticsnerds.blogspot.com.es/2016/02/a-cautionary-tale-of-bioinformatics.html
http://blogs.discovermagazine.com/loom/2012/07/19/the-mystery-of-the-missing-chromosome-with-a-special-guest-appearance-from-facebook-creationists/#.WL9GDH-P_1B (y varios artículos de continuación)
http://theleagueofreason.co.uk/viewtopic.php?f=8&t=12424
http://biology.stackexchange.com/questions/5558/chromosome-2-fusion
https://roohif.wordpress.com/
https://www.reddit.com/r/NaturalTheology/comments/2625uu/my_first_reply_to_jeffrey_tomkins/
https://www.reddit.com/r/junkscience/comments/3mtsto/the_chromosome_2_fusion_site_part_1_a_lack_of/
https://www.reddit.com/r/junkscience/comments/3n4vim/the_chromosome_2_fusion_site_part_2_the_fossil/
https://www.reddit.com/r/junkscience/comments/3ofwf8/human_chimp_similarity_take_2/
20# Lewis, C.S.. Apologética cristiana (1945). Publicado en: Lo eterno sin disimulo. Rialp, Madrid, 1999, p.19.


Que la ciencia sea incapaz de ofrecer la explicación última del cosmos, no quiere decir que tal explicación no pueda alcanzarse por otro camino.

Reina, Antonio del Corro y Cipriano de Valera, entre otros, se identificaron plenamente con los postulados evangélicos y se distanciaron de las enseñanzas de la Iglesia católica romana.

Introducimos dos conceptos muy importantes para el multimedia, aunque ciertamente se pueden aplicar a otras áreas también.

¿Qué es realmente lo que nuestra vida cristiana, ante la iglesia y ante la sociedad está mostrando?

Se llevan a cabo el Concordato de la Santa Sede y los Pactos con Estados Unidos de América.

No soy persona de anotar, lo que tal vez no pueda después cumplir; prefiero acabar y terminar mi año encomendándome al Señor de mi vida.

Como cristiano, Unamuno renuncia a la venganza. Sabe que en esos momentos no habrá paz, aquella que esperaba encontrar luchando con la palabra como espada.
 La década en resumen: teología, con José Hutter
La década en resumen: teología, con José Hutter
La conmemoración de la Reforma, las tensiones en torno a la interpretación bíblica de la sexualidad o el crecimiento de las iglesias en Asia o África son algunos de los temas de la década que analizamos.
 Intervalos: Disfruten de la luz
Intervalos: Disfruten de la luz
Estudiamos el fenómeno de la luz partiendo de varios detalles del milagro de la vista en Marcos 8:24, en el que Jesús nos ayuda a comprender nuestra necesidad de ver la realidad claramente.
 2020, año del Brexit
2020, año del Brexit
Causas del triunfo de Boris Johnson y del Brexit; y sus consecuencias para la Unión Europea y la agenda globalista. Una entrevista a César Vidal.
 7 Días 1x08: Irak, aborto el LatAm y el evangelio en el trabajo
7 Días 1x08: Irak, aborto el LatAm y el evangelio en el trabajo
Analizamos las noticias más relevantes de la semana.
 Min19: Infancia, familia e iglesias
Min19: Infancia, familia e iglesias
Algunas imágenes del primer congreso protestante sobre ministerios con la infancia y la familia, celebrado en Madrid.
 X Encuentro de Literatura Cristiana
X Encuentro de Literatura Cristiana
Algunas fotos de la entrega del Premio Jorge Borrow 2019 y de este encuentro de referencia, celebrado el sábado en la Facultad de Filología y en el Ayuntamiento de Salamanca. Fotos de MGala.
 Idea2019, en fotos
Idea2019, en fotos
Instantáneas del fin de semana de la Alianza Evangélica Española en Murcia, donde se desarrolló el programa con el lema ‘El poder transformador de lo pequeño’.
 Héroes: un padre extraordinario
Héroes: un padre extraordinario
José era alguien de una gran lealtad, la cual demostró con su actitud y acciones.
 Programa especial de Navidad en TVE
Programa especial de Navidad en TVE
Celebración de Navidad evangélica, desde la Iglesia Evangélica Bautista Buen Pastor, en Madrid.
 Primer Congreso sobre infancia y familia, primera ponencia
Primer Congreso sobre infancia y familia, primera ponencia
Madrid acoge el min19, donde ministerios evangélicos de toda España conversan sobre los desafíos de la infancia en el mundo actual.














Las opiniones vertidas por nuestros colaboradores se realizan a nivel personal, pudiendo coincidir o no con la postura de la dirección de Protestante Digital.



{kind=link}
{kind=link}
{kind=link}
Si quieres comentar o